
Ecophysiological characteristics of two planktonic desmid
species originating from trophically different lakes
Elly Spijkerman & Peter F.M. Coesel
Abstract
Introduction
Materials and methods
Results & Discussion
Published in Hydrobiologia
Two planktonic desmid species were compared in some of their ecophysiological characteristics. Staurastrum chaetoceras, well-known for its abundant occurrence in eutrophic lakes, showed a higher photosynthetic capacity and a higher maximum (intrinsic) growth rate than Cosmarium abbreviatum var. planctonicum, a taxon only encountered in oligo-mesotrophic habitats. The two taxa are comparable in cell size. When grown under a stringent continuous inorganic phosphorus (Pi) limitation, C. abbreviatum realized a higher growth rate, due to a higher affinity for the uptake of Pi, than S. chaetoceras. On the other hand, under those conditions, S. chaetoceras displayed a two times higher maximum Pi uptake rate (Vi,max). Regarding cellular alkaline phosphatase activity (hydrolysis of the organic P substrate MFP) C. abbreviatum showed both a higher affinity and maximum rate than S. chaetoceras.
In a way, these characteristics reflect the distribution pattern of the two species in the field. For in eutrophic lakes, during the summer algal bloom, species often have to compete for light as the growth limiting factor, whereas species occurring in oligo-mesotrophic lakes usually face permanently growth-limiting P concentrations. Since in eutrophic lakes during summer algal bloom dissolved inorganic P concentrations can also be low, the ability of phytoplankton to acquire Pi from short-lived pulses (e.g. excretion of P by zooplankton or fish) has to be considered an important additional characteristic in view of competition. Concerning the two desmid species under discussion, S. chaetoceras will have a competitive advantage when Pi is supplied in distinct pulses, due to its higher Vi,max values. On the other hand, C. abbreviatum possibly will be superior in competition for organic P substrates. In the species studied, different strategies were found to benefit optimally from the resource conditions inherent in the trophic state of their habitat.
The ability of a given phytoplankton species to sustain a population in an oligotrophic environment acquires ecophysiological characteristics different from those to be expected in a eutrophic habitat. When the summer species composition of trophically distinct hard-water lakes from the temperate zone are compared, specific differences in the growth and nutrient-uptake characteristics of the main species are anticipated (Hecky & Kilham 1974, Tilman 1977, Kilham & Tilman 1979). Conversely, nutrient uptake and growth kinetics may help to explain the distribution of algal species in the field. Cell size is found to be an important parameter determining ecophysiological parameters (Smith & Kalff 1982), but opposite opinions exist about the relation between predominant algal cell size and nutrient load of a lake. Smaller cells, with their high surface area-to-volume ratio, generally have high nutrient uptake affinities and thus would benefit more from a low nutrient load in a lake (e.g. Herbland et al. 1985, Watson & Kalff 1981, Wehr 1993). On the other hand it is said that nutrient-depleted waters favour the occurrence of large cell-sized phytoplankton species because of their lower maximum growth rate (µmax) (Maestrini & Bonin 1981), which would be of survival value (Lund 1965). This is inconsistent with the conclusion of Reynolds (1987) that species with similar cell size and cell shape demonstrate similar growth and survival strategies. Guillard et al. (1973) compared two clones of the diatom Thalassiosira pseudonana Hasle et Heimdal in their silicate-limited growth characteristics and found that the affinity of the clone from the oligotrophic site was higher and its µmax was significantly lower than the clone from the eutrophic site. The two planktonic desmid taxa used in the present study, Cosmarium abbreviatum Rac. var. planctonicum W. & G. S. West (characteristic of an oligo-mesotrophic environment) and Staurastrum chaetoceras (Schr.) G.M. Smith (peaking in eutrophic water bodies: Coesel 1994) are comparable in cell size. Possible differences in kinetic characteristics, caused by cellular surface-to-volume ratios are thus reduced to a minimum. Both species amass their maximum population densities in the summer period when algal biomass is relatively high and dissolved nutrient concentrations are low. The desmid taxa were compared in their maximum growth rate, growth under light or continuous phosphorus- (Pi) limited conditions, Pi-uptake rates and alkaline phosphatase activity as possible indicators for their pronounced distribution pattern in the field.
High maximum P-uptake rates and maximum growth rates enable a species to invade an environment with (temporary) high ambient nutrient concentrations (r-strategist). Low maximum growth rates and high affinity for P uptake reflect the survival strategy of a species growing under low nutrient conditions (K-strategist, Sommer 1981). During a summer bloom, low light intensities will benefit the growth of algal species with a high photosynthetic capacity. As for phosphorus, usually acting as the limiting nutrient, Hantke et al. (1996b) found that more than half of the total P content of a eutrophic lake was found in particles and the dissolved P fraction contained twice as much DOP (dissolved organic P) as SRP (soluble reactive P). Since it can be assumed that 60% of DOP consists of suitable substrates for phosphatases (Hantke et al. 1996b), alkaline phosphatase activity (APA) reflects the ability of algae to use organic P sources for growth.
The present paper is a concise review of previously presented findings about the growth and nutrient uptake kinetics of C. abbreviatum and S. chaetoceras, complemented with recently obtained data concerning alkaline phosphatase activities.
Algal isolates— The experiments were performed with Staurastrum chaetoceras, clone AO 36, isolated from the alkaline eutrophic Lake IJmeer (Berger & Sweers 1988) and Cosmarium abbreviatum var. planctonicum, clone AO 116, isolated from the alkaline, oligo- mesotrophic Lake Maarsseveen (I) (Swain et al. 1987). Characteristics of the lakes are listed in Table 7-I. Both clones were taken from the desmid collection in the Department of Aquatic Ecology, University of Amsterdam.
Culturing conditions in a temperature-light gradient— The species were batch-cultured in glass dishes with 50 ml Woods Hole medium. The growth rates at different combinations of temperature and incident irradiance were assessed in an apparatus that combines a temperature gradient with a light gradient at right angles. Algal cells were counted with a Coulter Multisizer. All experiments were performed under a light-dark cycle of 16:8. For more detailed information, see Coesel & Wardenaar (1990).
Culturing conditions in turbidostat and chemostat— The species were cultured at 20 ± 1 °C in 1 L continuous-flow culture vessels. The inflow medium for the light-limited cultures (turbidostat) contained 50 µM P, while for continuously Pi-limited cultures (chemostat) it contained only 5 µM P. For details of the continuous-flow device and turbidostat experiments, see Coesel & Wardenaar (1994), of the composition of the P-limited medium, see chapter 2. Average irradiance in the culture vessel was calculated according to Van Liere et al. (1978). Illumination of the uni-algal cultures was continuous. Cultures were not axenic but bacterial biomass (estimated by acridine staining and counting under a fluorescence microscope) was negligible (<1% of algal biomass). Chemostat cultures were considered to be in a steady state (dilution rate D = growth rate µ) when algal biomass varied less than 5% over at least 2 renewals of the culture vessel volume. For details about calculations of chemostat culture parameters, see chapter 2.
P-uptake experiments— Uptake experiments were performed with culture material from steady-state conditions, which was two-fold diluted with P-free culture medium and pulsed with different inorganic phosphorus (Pi) concentrations containing 32P as described in chapter 2. Initial Pi concentrations ranged from 0.5 to 30 µM. Cellular 32P contents were determined at t = 0, 30, 60 and 300 s, and initial uptake rates (V) for every initial Pi concentration were calculated from linear regression, following Riegman & Mur (1984b). By curve fitting to the Michaelis-Menten equation
V = Vi,max * (Pi / (Pi + Km))
maximum uptake rate (Vi,max) and half-saturation constant for uptake (Km) were computed.
Table 7-I. Some characteristics of Lake Maarsseveen I (Lingeman et al. 1987, Swain et al. 1987) and Lake IJmeer (Berger & Sweers 1988, unpublished data of the Ministry of Public Works, directorate of IJsselmeer area). The IJmeer is part of the IJsselmeer.
|
|
Lake Maarsseveen I |
Lake IJmeer |
|
location (Lat., Long.) |
5°04'E - 52°09'N |
5°04'E - 52°21'N |
|
surface area (ha) |
70 |
4500 |
|
max. lenght (m) |
1,760 |
20,000 |
|
max. depth (m) |
30.8 |
25 ( in local pits) |
|
mean depth (m) |
12.1 |
4.5 |
|
average period of stratification |
may - october |
no stratification periodin 90% of the area. |
|
average depth of thermocline (m) |
8 - 13 |
- |
|
range of vertical extinction coeff. (m-1) |
0.4 - 0.6 |
1.0 - 4.6 |
|
range of Secchi disc transparencies (m) |
3.5 - 8.0 |
0.2 - 0.9 |
|
P-total (mg L-1) |
0.009 - 0.012 |
0.07 - 0.32 |
|
N-total (mg L-1) |
0.1 - 0.6 |
1.1 - 2.0 |
|
conductivity (µS cm-1) |
350 - 500 |
600 - 900 |
|
pH |
7.5 - 8.3 |
7.8 - 9.2 |
|
Chl-a maximum (µg L-1) |
4 |
47 |
|
phytoplankton characterisation |
Chrysophyceae |
Chlorococcales |
Alkaline phosphatase activity— The alkaline phosphatase activity (APA) was determined on cells from continuous-flow cultures with an inflow medium free from P, adapted to a pulse of 10 µM Pi every two weeks (D = 0.007 h-1). Starved cells were harvested right before the pulse was given to the culture, washed and resuspended in P-free Woods Hole medium without Fe-EDTA (see Jansson et al. 1988). The medium was brought to a pH 8.5 to optimize the enzyme activity. APA was measured by adding 3-0-MFP (methylfluoresceinphosphate) to the cells and measuring the release of MF (methylfluorescein) in time on a recorder connected to a Perklin Elmer fluorimeter (Perry 1972). The initial concentrations of MFP ranged from 0.05 to 4.5 µM. The determination (in duplicate) was repeated 10 weeks later in another sample from the continuous-flow culture in a comparable starved condition.
One of the main results obtained is the higher intrinsic growth rate of Staurastrum chaetoceras as compared to Cosmarium abbreviatum, this in spite of the fact that the two species are comparable in cell size. As is shown in Figure 7-1, the difference in growth rate in favour of S. chaetoceras increases with increasing light intensity and temperature. Actually, the maximum light intensity as offered by the light-temperature apparatus appeared to be still suboptimal for S. chaetoceras. Growth rate of the latter species could be raised to µ > 0.042 h-1 providing higher incident irradiance (not shown).
Figure 7-1. Growth response (µ, h-1) of Cosmarium abbreviatum and Staurastrum chaetoceras tested in the light-temperature gradient apparatus. (After Coesel & Wardenaar 1990)
The higher growth response to light intensity in S. chaetoceras as compared to C. abbreviatum (Figure 7-1) was confirmed in the turbidostat experiments at a fixed temperature of 20 °C (Figure 7-2) and could be related to a higher photosynthetic capacity in S. chaetoceras (Coesel & Wardenaar 1994). From these data, it is likely that S. chaetoceras will have a relatively high competitive ability in shallow lakes where light climate and temperature in summer are favourable to realize near µmax values, provided of course, that there are enough nutrients available.
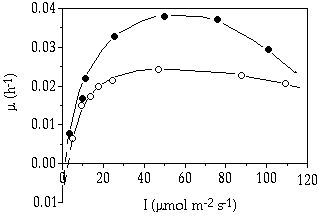
Figure 7-2. Growth rate (µ, h-1)
in relation to irradiance (I) in Cosmarium abbreviatum (![]() ) and Staurastrum chaetoceras (
) and Staurastrum chaetoceras (![]() ) grown in continuously illuminated turbidostat cultures. (After Coesel &
Wardenaar 1994)
) grown in continuously illuminated turbidostat cultures. (After Coesel &
Wardenaar 1994)
In this connection it is important to be able to benefit from temporary high concentrations of essential nutrients. Indeed, the maximum uptake rate (Vi,max) for phosphate (usually a growth limiting nutrient in fresh waters) in S. chaetoceras appeared to be distinctly higher than that in C. abbreviatum (Table 7-II). These characteristics (high Vi,max and µmax) can be considered adaptations to a dynamic environment (Zevenboom 1986).
Table 7-II. Maximum uptake rates (Vi,max) in µmol P mg protein-1 h-1, half saturation constants (Km) in µM P and affinities (Vi,max/Km). Mean values ± standard deviation of C. abbreviatum and S. chaetoceras at different steady-state growth rates (µ). Number of replicates in parentheses. (After chapter 3)
|
|
Cosmarium abbreviatum |
Staurastrum chaetoceras |
||||||
|
µ (h-1) |
Vi,max |
Km |
Vi,max/Km |
n |
Vi,max |
Km |
Vi,max/Km |
n |
|
0.003 |
0.45 ± 0.10 |
1.54 ± 0.76 |
0.33 ± 0.14 |
(4) |
|
|
|
|
|
0.005 |
0.47 |
1.60 |
0.30 |
(1) |
0.81 ± 0.22 |
4.09 ± 1.27 |
0.20 ± 0.05 |
(6) |
|
0.007 |
" 0.34 ± 0.01 |
1.51 ± 0.84 |
0.27 ± 0.14 |
(2) |
|
|
|
|
|
0.010 |
0.40 ± 0.11 |
1.29 ± 0.54 |
0.33 ± 0.07 |
(3) |
0.91 ± 0.22 |
6.44 ± 1.56 |
0.14 ± 0.02 |
(3) |
|
0.015 |
0.27 ± 0.10 |
1.97 ± 1.02 |
0.17 ± 0.12 |
(3) |
0.95 ± 0.09 |
5.80 ± 1.08 |
0.17 ± 0.03 |
(4) |
|
0.020 |
0.65 ± 0.10 |
12.44 ± 1.76 |
0.05 ± 0.01 |
(3) |
0.98 ± 0.21 |
6.95 ± 2.10 |
0.15 ± 0.02 |
(3) |
|
0.030 |
|
|
|
|
0.57 ± 0.12 |
2.95 ± 0.72 |
0.20 ± 0.02 |
(3) |
|
0.035 |
|
|
|
|
0.53 ± 0.34 |
6.94 ± 2.61 |
0.07 ± 0.04 |
(3) |
Growth responses under a continuous P limitation (Figure 7-3) show that, at stringent P limited conditions, C. abbreviatum can realise a higher growth rate, due to a higher affinity for P, than S. chaetoceras (Table 7-II).
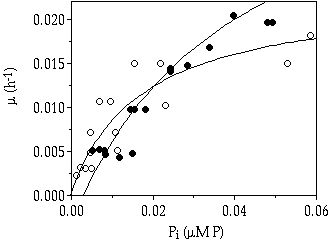
Figure 7-3. Growth rate (µ, h-1)
in relation to the inorganic phosphorus concentration in the culture vessel
(Pi) of Cosmarium abbreviatum
(![]() ) and Staurastrum chaetoceras
(
) and Staurastrum chaetoceras
(![]() ). Curves fitted to the Monod model.
(From chapter 3)
). Curves fitted to the Monod model.
(From chapter 3)
The results from Figure 7-2 and 7-3 concerning two particular growth rates, i.e. µ = 0.007 h-1 and µ = 0.015 h-1 are combined in Figure 7-4. This figure shows isoclines that indicate the minimum resource levels needed to sustain a rate of growth adequate to exceed the loss rate imposed (in steady-state continuous flow culture: loss rate = dilution rate = growth rate). This rendering is based on a general concept by Phillips (1973), which concept was extended by Taylor & Williams (1975) for growth of micro-organisms under substrate limitation. The theory in question has been worked out in more detail and applied for phytoplankton studies by Tilman (1980) and Tilman et al. (1982). Huisman & Weissing (1994, 1995) were the first to extend this theory to a combination of light and nutrient limitation.
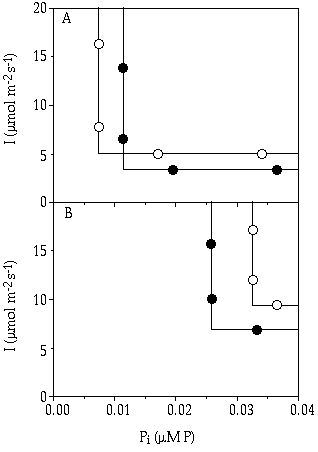
Figure 7-4. Growth isoclines of Cosmarium abbreviatum
(—![]() —) and Staurastrum chaetoceras
(—
—) and Staurastrum chaetoceras
(—![]() —) at a loss rate of (A) D = 0.007
h-1 and (B) D = 0.015 h-1
indicating their competitive ability for light (I) and phosphorus (Pi)
limiting conditions.
—) at a loss rate of (A) D = 0.007
h-1 and (B) D = 0.015 h-1
indicating their competitive ability for light (I) and phosphorus (Pi)
limiting conditions.
This is more complicated in view of the light gradient caused by shading and mutual interference of light- and nutrient-limited growth responses. Healey (1985) found that the internal concentration of the limiting nutrient of the cyanobacterium Synechococcus linearis (Naeg.) Kom. increased with decreasing irradiance. This effect was the most pronounced with nitrate and also found for Pseudoanabaena sp. (see also Rhee & Gotham 1981). Knoechel & deNoyelles (1980) found that Dinobryon sertularia from natural phytoplankton responded to changes in both light and P supply rates. Because of these interactions, the isoclines in Figure 7-4 most probably have round corners but, because no data concerning this situation have been obtained, only straight, right-angled isoclines are inserted here, for simplicity. Huisman & Weissing (1995) showed in their model that it is not just the ratio of nutrient supply to light supply, but also the absolute supply rate of the light that determines the outcome of competition. This is a fundamental difference with the nutrient ratio theory of Tilman (1982). Despite all simplifications applied to the isoclines in Figure 7-4, it is shown that at a low loss rate (in ecological terms, a weakly dynamic environment) C. abbreviatum is better adapted to utilize low P concentrations, whereas S. chaetoceras is in advantage at low light intensities (Figure 7-4a). In contrast to that, at higher loss rates (as met with in a more dynamic environment) C. abbreviatum has a disadvantage at any combination of irradiance and phosphate (Figure 7-4b).
Another distinct difference between the two desmid species was found in their cellular-bound alkaline phosphatase activities (Figure 7-5). Both the maximum hydrolysation rate (APAmax) and the affinity to the substrate were pronouncedly higher in C. abbreviatum. This was not due to bacterial activity, because the cells were washed by centrifugation.
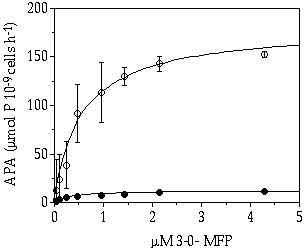
Figure 7-5. Alkaline phosphatase activity (APA) of Cosmarium
abbreviatum (![]() ) and Staurastrum
chaetoceras (
) and Staurastrum
chaetoceras (![]() ) from starved continuous
flow cultures in relation to the concentration of added 3-0-methylfluoresceinphosphate
(MFP).
) from starved continuous
flow cultures in relation to the concentration of added 3-0-methylfluoresceinphosphate
(MFP).
From data presented in Figure 7-5 and Table 7-II it can be calculated that C. abbreviatum, with its cell-bound phosphatases can free more than enough Pi from an organic P substrate (APAmax), to meet its maximum Pi uptake rate (Vi,max). This in contrast to S. chaetoceras where APAmax was much lower and Vi,max much higher than in C. abbreviatum. It is well-known that phosphatases are excreted by different planktonic organisms (e.g. Kuenzler & Perras 1965, Berman 1970) and our data suggest that C. abbreviatum may sequester enough Pi to satisfy its own requirements by hydrolysation of organic P compounds via cellular APA, whereas S. chaetoceras will benefit more from temporary high concentrations of Pi released by dissolved AP (supplied by other planktonic organisms). Our findings can not be directly linked to fieldstudies on APA. Hantke et al. (1996b) found increasing total phosphatase activities with higher eutrophication level, and the algal fraction in general contributed less than half of the total APA. In the oligotrophic lake they studied 50% of the total hydrolysis was realized by the dissolved fraction and only 13% by phytoplankton (Hantke et al. 1996b). From the data of Hantke et al. (1996a & b) referring to the upper layer in the summer period it can be calculated that the fraction from the oligotrophic lake containing the algal cells (>0.1 µm) had a higher APAmax per unit of chlorophyll-a than the corresponding fraction from the meso- and eutrophic lakes, while the affinity of the phytoplankton fraction (APAmax/Km) was not different between the lakes. This is partly in agreement with our data that C. abbreviatum has a higher capacity for the hydrolysis of organic phosphates.
A distinct morphological difference between C. abbreviatum and S. chaetoceras (next to the difference in cell shape) is the fact that C. abbreviatum has a mucilage sheath around the cell which is maintained under all possible physiological conditions (Coesel 1994). The higher APA of C. abbreviatum could be linked to this phenomenon if the mucilage sheath surrounding the cells of C. abbreviatum would act as a storage for AP, but we were unable to demonstrate this by shaking or sonification, because these treatments released the cell-bound APA of S. chaetoceras as well. We failed to detect bacteria in the sheath as a possible source for APA, so have to conclude that the algal cells themselves were responsible for the high enzyme production. That the algal cell sheath might act as a primary or secundary location for AP was also suggested by Doonan & Jensen (1977), but they could not prove it either.
The observed species specific differences in the ecophysiological characteristics fit very well to the origin of the algal strains used. For Staurastrum chaetoceras, characterized by a relatively high photosynthetic capacity, high maximum P-uptake rate and high maximum growth rate, was isolated from a eutrophic lake, where, in summer time, light climate and dissolved nutrient concentrations are usually highly variable. In contrast, Cosmarium abbreviatum, marked by a relatively high affinity for inorganic P uptake and a high alkaline phosphatase activity originated from an oligo-mesotrophic clearwater lake, where phosphorus is supposed to be the main algal growth limiting factor. So, although cell size may be a major factor in predicting competition between phytoplankton species for a limiting nutrient, the potential variation in uptake parameters among species of comparable size should not be underestimated.